The advent of CRISPR-Cas9-based genome editing offered a convenient alternative to existing complex techniques such as zinc-finger nucleases. Using CRISPR-Cas9, researchers can edit DNA sequences in the genome with utmost precision. The technique has resulted in easier ways to insert or delete genes, tweak single base pairs in DNA, and has garnered unforeseen adoption in the research community.
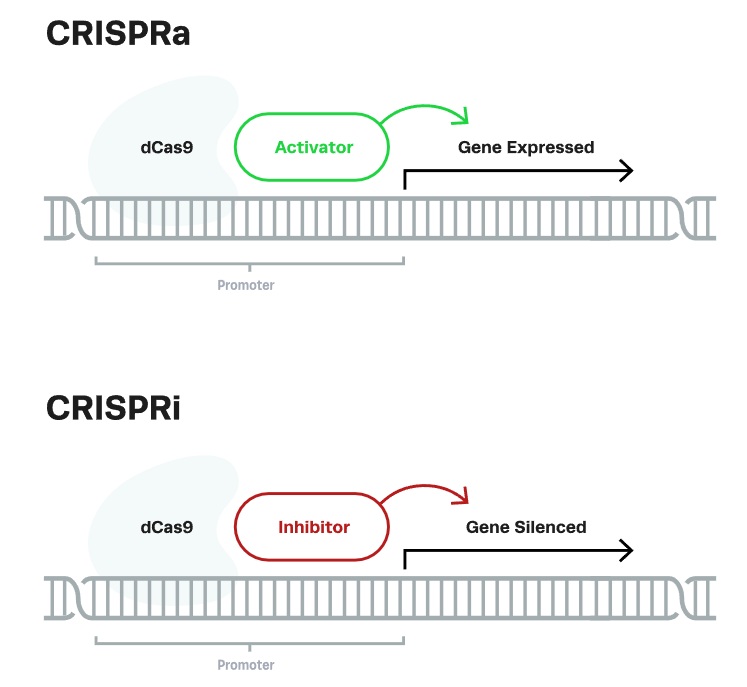
But CRISPR technology has the power to go beyond gene editing — a slight tweak in the CRISPR-Cas9 duo turns it into a gene-specific regulator, adding the functionality of a light dimmer to an already formidable on-off switch. The sub-applications of CRISPR that render the brightening and dimming are called CRISPRa and CRISPRi, short for CRISPR activation and interference, respectively.
CRISPR activation or CRISPRa is a variant of CRISPR in which a catalytically dead (d) Cas9 is fused with a transcriptional effector to modulate target gene expression. Once the guide RNA navigates to the genome locus along with the effector arm, the dCas9 is unable to make a cut, and instead, the effector activates the downstream gene expression.
CRISPR interference or CRISPRi is also a variant of CRISPR in which a catalytically dead (d) Cas9 is fused with a transcriptional effector to modulate target gene expression. However, in CRISPRi, when guide RNA navigates to the genome locus along with the effector arm, it represses the downstream gene expression instead of activating it.
While CRISPRa/i modulate gene expression transiently, gene knockouts disrupt the gene function permanently. While useful to screen weak phenotypes that can be masked by residual low-level gene expression, knockouts are not suited to studying essential genes, because deleting such genes results in cellular toxicity. In addition, gene regulation is useful to study pharmacotherapy because drugs often reduce gene expression, rarely do they wipe out a gene completely. Therefore, knocking down a gene partially can mimic drug action better than a knockout.
Both CRISPRa and CRISPRi work with genes at the transcriptional level. Playing with the levels at which a target gene is expressed can help answer complex questions about gene function in cell signaling cascades. By sliding gene expression up and down, scientists can test the robustness and sensitivity of biological systems to those small changes.
CRISPRa allows scientists the unprecedented ability to increase the expression of genes in their native context. This is in sharp contrast to other gain-of-function techniques such as plasmid and lentiviral-based open reading frames, or ORFs, that can primarily drive exogenous gene expression. CRISPRa also enables the overexpression of large transcripts for which ORF-based methods do not work as well.
CRISPRi is complementary to RNA interference or RNAi, but is advantageous in having fewer sequence-specific off-target effects, and in being able to modulate both coding as well as non-coding genes.
The CRISPR-Cas9 system makes use of an engineered single guide RNA (sgRNA) to home in the Cas9 nuclease to a specific locus in the genome. Cas9 introduces a double-stranded cut at the locus. The DNA damage activates a routine but error-prone process to repair double-stranded breaks, which introduces mutations and results in the complete loss-of-function of the target gene.
In prokaryotes, replacing Cas9 with a catalytically inactive version, dCas9, and targeting it to a promoter region can inhibit downstream gene expression because dCas9 sterically hinders transcriptional machinery. In mammalian cells, dCas9 alone achieves modest repression (60-80%), therefore the CRISPRi system includes dCas9 fused with a repressor domain such as Kruppel associated box, or KRAB, which can help silence genes in an inducible, reversible, and non-toxic manner.
Similarly, CRISPRa uses dCas9 fusions to activate gene expression. dCas9 fused to transcriptional activators such as VP64 and p65 can be targeted to promoter and enhancer regions, which results in higher than usual transcription of the target gene. To enhance gene activation in mammalian cells, dCas9 fusions that recruit multiple activator domains have also been developed, including a protein scaffold called the SunTag system, a sequence containing many copies of the activator recruitment domain of general control protein (GCN4).
A fundamental requirement of CRISPR-based gene regulation is for the sgRNA-dCas9 system to be able to find and bind to the promoter region of the target gene.
The first step is to design the sgRNA complementary to the promoter region or transcriptional start site. However, this is a challenge because start sites are not always well annotated in the genome. In addition, sometimes cryptic or alternative promoters located far upstream of the target gene can also come into play, further complicating design considerations. Even if the sgRNA is designed to find the promoter correctly, the promoter site may already be occupied by other protein factors, and therefore be inaccessible for the guide RNA-dCas9 system to bind.
In order to design the perfect sgRNA with the ability to regulate each gene, researchers have employed systematic screens at the genome-scale — by pooling a sgRNA library followed by next-generation sequencing, guide RNA specificity and efficacy can be evaluated for each gene. For example, if the expression of a certain gene went up or down, the specific sgRNA that drove it is considered efficacious. Using these data, researchers have built design algorithms to create lists of top sgRNA sequences for each gene in human and mouse genomes.
Getting from the perfect sgRNA sequence to making it can involve either plasmid-based expression or synthetic production. Cloning using a plasmid vector is one of the earliest methods to produce sgRNA, however, it is also time-consuming, and prone to off-target effects. Instead, synthetic sgRNA production offers a faster and more accurate method to get to high CRISPR editing efficiencies.
In order to further optimize CRISPR-based gene regulation, researchers are also engineering dCas9 to be less toxic by reducing its non-specific binding to NGG sequences (where N can be any nucleotide base).
CRISPRa and CRISPRi are handy tools to manipulate gene function, adding significantly to the pre-existing arsenal of genetic techniques. These tools lend themselves well to screens for functional genomics — by using a genome-wide library of guide RNAs, researchers can slide gene expression up or down in an unbiased manner, and screen for genes that affect growth and proliferation, or genes that mediate sensitivity to a selective pressure such as a drug or a toxin. Screens using CRISPRa and CRISPRi have been employed to study growth-driving genes in leukemia cells, and genes that mediate resistance to a BRAF inhibitor in melanoma cells. These techniques have found use in both basic and applied life sciences research.
Resistance to chemotherapy plays a significant role in cancer mortality. Bester et al used CRISPRa to identify genetic units that mediate resistance to chemotherapy, specifically to cytarabine, the cornerstone treatment for acute myeloid leukemia. Using the technique, they screened 14,701 long non-coding RNA genes, and identified novel cell-cycle, survival/apoptosis, and cancer signaling genes of therapeutic relevance.
Wangensteen et al adapted the CRISPRa system for use in vivo to assess whether changing endogenous gene expression levels can lead to functional changes in the native environment of the liver. The authors used CRISPRa screening to identify protein-coding genes that drive hepatocyte proliferation and tumorigenesis in an in vivo live mouse model of liver injury and repopulation. The authors reported a significantly enriched proto‐oncogene and development of hepatocellular carcinoma, both indicative of increased cell division and cancer initiation events.
In addition to using cancer cell lines, researchers have demonstrated the use of CRISPRi screens in induced pluripotent stem cells (iPSCs). For example, Tian et al used CRISPRi in human iPSC-derived neurons to conduct a growth screen and uncovered genes essential for neurons, but not iPSCs or cancer cells.
Taking CRISPRi to a new species where traditional genetic manipulation is challenging, Walker et al reported adapting CRISPRi to probe gene function in the malaria parasite Plasmodium yoelii, which will ultimately help control its transmission.
Overall, the CRISPR suite of technologies are powerful tools to probe gene function in multiple biological pathways and across disease areas, and the flexibility of the system offers endless opportunities.
https://doi.org/10.1021/acschembio.7b00657
https://doi.org/10.7554/eLife.19760.001
https://doi.org/10.1016/j.cell.2018.03.052
https://doi.org/10.1002/hep.29626
This protocol describes the use of limiting dilution to isolate single cells (clones) from a CRISPR edited population, the expansion of clonal populations, and screening to identify clones with the...
Thank you for choosing SpCas9 nuclease protein and gRNAs for your CRISPR experiment! SpCas9 has been used for diverse genome editing applications, making it the gold standard in CRISPR gene...
Learn more about clinical applications of CRISPR and how Synthego can support you from early-phase research, through process development, and into the clinic, highlighting our new sgRNA GMP manufacturing capabilities.