The CRISPR system evolved as a bacterial defense mechanism against viruses. Basically, upon encountering a viral infection, the bacterium uses this machinery called Cas, short for CRISPR-associated protein, to chop up the invading viral DNA. This fragment is then stored as genetic memory between repeating palindromic repeats called Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) to detect and combat future infections.
Once the underlying mechanism of CRISPR was discovered, researchers programmed this system to extend beyond bacteria and manipulate the genome of eukaryotic cells. CRISPR has since emerged as a powerful tool for genome engineering.
The simplicity of the system is one of its most valuable features for researchers. The CRISPR system comprises of two basic components: the guide RNA and the Cas nuclease. The guide RNA is complementary to the specific target DNA sequence, while the nuclease acts like a pair of molecular scissors that actually cleave the target DNA. Due to CRISPR's simplicity, researchers and scientists are using CRISPR across various applications including agriculture, diagnostics, biofuels, and most notably, cell and gene therapies for the treatment of disease in the clinic.
Cas9 is currently the most widely known nuclease in CRISPR experiments, specifically the Cas9 variant isolated from the bacterium Streptococcus pyogenes (SpCas9). SpCas9 nucleases require both crisprRNA (crRNA) and tracrRNA; as a complex, these two components are referred to as guide RNA (gRNA) or sometimes single guide RNA (sgRNA). When the gRNA directs Cas9 to the target DNA sequence, Cas9 additionally requires a short stretch of nucleotides, termed the protospacer adjacent motif (PAM) sequence, downstream of the target sequence for its cutting action. (Note, the PAM sequence is not part of the gRNA). Once Cas9 recognizes the PAM sequence (5’-NGG-3’ in the case of SpCas9, where N is any nucleotide), it creates a double-strand break (DSB) at the target locus. Cas9 activity is a collective effort of two parts of the protein: the recognition lobe and the nuclease lobe. One lobe recognizes the complementary DNA sequence, and the other has nuclease domains that cleave the DNA.
Although Cas9 is the most widely CRISPR variant used in genome engineering experiments, it does have certain limitations. First, its complementarity specifications are not very stringent - it can recognize other PAM sequences, like 5’-NAG-3’and 5’-NGA-3’, which results in off-target editing. This means it can sometimes cut non-specific DNA sequences. These off-target effects could cause detrimental effects in the cell and are especially concerning for the application of this method in humans. Second, its relatively large size makes it difficult to deliver inside cells using common viral vectors like adeno-associated viruses (AAVs). Lastly, the stringent requirement for the PAM sequence limits the target DNA that can be manipulated using this method.
Over the past few years, several CRISPR nuclease variants have been developed to overcome these issues. What are the different nuclease variants? Are these better than Cas9, and if so, how? Here, we answer these questions by summarizing the different nucleases available for a CRISPR experiment.

CRISPR has quickly become a standard laboratory tool for gene editing. As the adoption of CRISPR accelerates worldwide, up-to-date knowledge of the basics of CRISPR is essential for anyone in the field. From target identification studies to the recent breakthroughs in clinical trials, CRISPR is enabling scientists to unlock the power of the genome.
Download our CRISPR 101 eBook today to stay up to date on all your CRISPR basics and get the best results in your CRISPR experiments!
Although SpCas9 is the most popular nuclease, Cas9 can also be isolated from different species of bacteria. The Cas9 nucleases from different bacteria mainly differ in the PAM sequence that they require for cleavage. However, some of them also possess other differences. Let’s take a look at some of the naturally occurring Cas9 nuclease variants.
Staphylococcus aureus Cas9 (SaCas9) is an increasingly popular Cas nuclease. At just 1053 amino acids in length, it’s about 1 kb smaller than the traditional SpCas9. This means that it can be easily packaged into viral vectors, like the FDA-approved Adeno-associated viruses (or AAVs for short), making it ideal for clinical applications. SaCas9 recognizes a 3’-NNGRRT PAM sequence, generates blunt-end double-strand breaks (DSBs), and can be used in a paired nickase approach.
SaCas9’s versatility, precision, and small size have been utilized in a variety of clinical and research applications. For example, a 2020 paper published in Science Advances used SaCas9 to edit specific neuronal populations in rat brains for the study of neural circuitry. Similarly, SaCas9 has been used to generate cell-type-specific mutations in the nervous system of mice, enabling circuit-specific gene function research.
Being able to target specific cell populations, organs, or tissues is extremely relevant to gene therapies in human subjects. In a study published in Frontiers in Microbiology, the authors reported delivering SaCas9 via AAV8 with liver-specific promoters to inhibit the replication of the hepatitis B virus (HBV) both in vitro and in vivo. This treatment restricted the gene editing components to hepatocytes, reducing the potential for adverse events, which are a major concern in clinical studies.
This powerful nuclease was also used to investigate the role of the myostatin gene in muscular atrophy and sarcopenia. The pre-clinical study found that SaCas9-mediated knockout of the myostatin gene in a mouse model resulted in an increase in body weight, improved muscle cell differentiation, and prevention of muscular atrophy. This experimental treatment has significant therapeutic potential for age-related sarcopenia.
Like SpCas9, SaCas9 has also been engineered for a broader range of applications. A 2019 study engineered SaCas9-HF, a high-fidelity variant with genome-wide activity in human cells with no reduction in on-target editing efficiency. Another engineered variant, KKHSaCas9, recognizes a 5’-NNGRRT-3’ PAM, and has a 2-4x broader range of target sites in the human genome than the wildtype SaCas9.
This nuclease has been used to edit many plant genomes, including tobacco, potato, and rice. Many of the gene editing studies in agriculturally important plant species aim to improve stress tolerance and pathogen resistance. SaCas9 has also been used to study plant-pathogen interactions in the model organism Nicotiana benthamiana.
A 2019 study that compared the efficiency and specificity of various Cas nucleases for gene editing applications in plants found that SaCas9 was the most efficient at generating indels.
SaCas9 has also been adapted for use in adenine base editing (ABE). A 2020 paper in Nature Communications described microABE1744, an engineered SaCas9 variant with significantly improved on-target editing compared to other nucleases, with a reduced RNA off-target footprint. This variant is also one of the smallest Cas9-ABEs available that can be delivered with AAVs.
However you are using SaCas9, Synthego can provide you with the high-quality synthetic guide RNA you need for your SaCas9 gene editing applications.
Researchers have so far used Cas9 variants from Streptococcus thermophilus (StCas9), Neisseria meningitidis (NmCas9), Francisella novicida (FnCas9), and Campylobacter jejuni (CjCas9), to name a few, in CRISPR experiments.
Another Cas9 variant was isolated from the bacteria Streptococcus canis (ScCas9). The sequence of the ScCas9 nuclease bears striking similarity (89.2% sequence homology) to that of SpCas9, but differs in one important aspect. It requires a less stringent PAM sequence—5′-NNG-3′ (instead of 5′-NGG-3′ for SpCas9)— for its activity, largely expanding the genomic target for CRISPR editing. Also, a Cas9 variant was identified in Staphylococcus auricularis, SauriCas9, which recognizes a 5’-NNGG-3’ PAM sequence, has high editing activity, and is small enough for delivery in AAVs.
Alternative Cas nuclease variants found in nature may confer some advantages over SpCas9, but most issues persist. Researchers have therefore engineered novel nuclease variants that cater to specific experimental needs, rather than relying on natural nucleases.
Utilizing their HG-PRECISE platform, HuidaGene Therapeutics engineered hfCas12Max from Cas12i of the Cas12 type V CRISPR-Cas nuclease family. This engineering resulted in hfCas12Max having enhanced gene editing capabilities while reducing unwanted off-target editing - making hfCas12Max a high-fidelity nuclease. Although engineered, hfCas12Max produces staggered-end cuts like other Cas12 nucleases. In addition to its high fidelity, hfCas12Max has a broader PAM sequence recognition profile (5’ - TN) that enables scientists to target regions of the genome that other common Cas nuclease cannot.
What helps make hfCas12Max stand out as a potential nuclease to use in CRISPR-based therapeutic development is its small size. Being only 1080 amino acids and requiring only the crRNA which is shorter in length, hfCas12Max nuclease and gRNA could be packaged into lipid nanoparticles (LNPs) and AAV delivery systems. Currently, HuidaGene Therapeutics has developed a CRISPR-based therapeutic using hfCas12Max to treat Duchenne muscular dystrophy This drug is referred to as HG302 in their clinical pipeline. The next generation of CRISPR-based therapeutics could potentially use hfCas12Max.

Explore Synthego's high-fidelity novel nuclease, hfCas12Max and how it can accelerate the development of CRISPR-based therapeutics.
Developed by AstraZeneca, the eSpot-ON nuclease is an engineered Cas9 variant originally found in the genome of the bacterium Parasutterella secunda. The wild-type PsCas9 nuclease was highly active but lacked the high fidelity critical for the development of CRISPR medicines. By testing a variety of mutations to the RuvC, wedge [WED], and [PAM-interacting] domains of the nuclease, scientists created a superior engineered PsCas9 (published as ePsCas9) variant. Some engineered high-fidelity SpCas9 nucleases typically come with the trade-off of having lower on-target editing, but ePsCas9 achieves exceptionally low off-target editing while retaining its robust on-target activity.
Commercially available as eSpOT-ON in both recombinant protein and mRNA format, ePsCas9 nuclease is only one part of this high-fidelity CRISPR therapeutic solution. The optimized gRNA for eSpOT-ON is reduced in size, increasing its stability and supporting the interaction between nuclease and guide to enhance editing efficiencies. The eSpOT-ON nuclease recognizes the same PAM sequence as SpCas9 (NGG) but creates sticky-end DSBs, leaving three-nucleotide 5’ overhangs. This not only promotes gene insertion via HDR, but also reduces the likelihood of chromosomal translocations, which are one of the key safety concerns associated with gene therapies.
The team at AstraZeneca showed the promising value of eSpOT-ON for CRISPR therapies to treat rare diseases by demonstrating its high level of gene editing when delivered via LNP to a clinically relevant liver target in mice. For CRISPR scientists developing genomic medicines, eSpOT-ON could provide a good balance between safety and efficiency.
The fidelity of the CRISPR system is directly related to the tolerance of DNA mismatches. The possibility of Cas9 cutting DNA at off-target sites is an important safety concern, especially for clinical applications of CRISPR in the future.
To overcome this issue, researchers developed variants by mutating the catalytic domains of Cas9, while retaining the specificity, such that the variants nicked a single DNA strand at the desired locus, instead of creating a double-strand break.
As a nick in the DNA is repaired with higher fidelity than a DSB, two nickases targeting opposite strands of DNA, each with a different guide RNA, could be used to create a double-strand break with high fidelity. Using such an approach, Cho et al. reported generating a relatively large 1 kb deletion in human cells with the off-target effects reduced to below the limit of detection.
Although nickases show high specificity, they rely on two guide RNAs to reach the target sites, thereby reducing the number of potential target sites in the genome. Alternatives to the wild type SpCas9 have now been engineered, including high fidelity variants that have a weakened DNA phosphate backbone interaction with the nuclease. The resulting SpCas9-HF1 nuclease displays genome-wide specificity and undetectable off-target effects, all with a single guide RNA (sgRNA).
Feng Zhang and colleagues developed an enhanced nuclease (eSpCas9) with higher fidelity than standard Cas9. Based on the crystal structure and other studies of the Cas9 nuclease, they hypothesized that when the guide RNA reaches its target DNA sequence, the double-stranded DNA strands separate as the RNA binds competitively to its target DNA strand. The nuclease has a groove lined with positive charges hypothesized to stabilize the negatively charged DNA strand. The researchers made changes to the amino acid residues in this groove to weaken interactions between the eSpCas9 and guide RNAs with non-exact complementarity with the target DNA sequence. This engineered Cas9 variant demonstrated improved specificity and lower off-target editing rates.
Armed with this knowledge and further single-molecule FRET experiments, Jennifer Doudna’s group found that the nuclease activity occurs after conformational changes in the HNH nuclease domain. This domain is controlled by the Rec3 domain, which is part of the recognition lobe of Cas9 that senses mismatches. Exploiting the natural conformational threshold, the researchers mutated Rec3 to make it more stringent and avoid cutting of mismatched DNA sequences. They named this version hyper-accurate Cas9 (HypaCas9), which improves proofreading before cleavage, and thus drastically reduces off-target cleavage. This variant has been shown to accurately edit mouse zygotes.
In 2014, David Liu’s team came up with a novel idea to deal with the off-target effects of Cas9: combining the DNA recognition of CRISPR with the specificity of another active nuclease FokI. This nuclease is known to cut only after dimerization, which is more difficult to occur at off-target sites than monomeric activity, thus resulting in enhanced specificity. As the activity depends on two guide RNAs binding to their complementary DNA targets, the number of sites targeted is lower than those targeted with active Cas9 nuclease.
A common limitation of all Cas9-based CRISPR variants is the requirement for specific PAM sequences next to the site of cleavage. Consequently, their nuclease activity is limited to about 1 in 16 sites in the genome. Recently, researchers at Harvard and MIT published a paper describing a newly engineered CRISPR enzyme called xCas9 to overcome this limitation. The xCas9 nuclease recognizes a broad range of PAM sequences, increasing the target sites to 1 in 4 in the genome. Remarkably, this CRISPR variant also exhibited lower off-target rates than the commonly used SpCas9. This shows great promise to expand the target areas of CRISPR technology. A study published in Science in 2020, developed novel Cas9 nucleases with altered PAM sequence specificities. The study developed two key variants: SpG, which has an expanded target range of PAM sequences, and SpRY, which can target almost all PAM sequences.
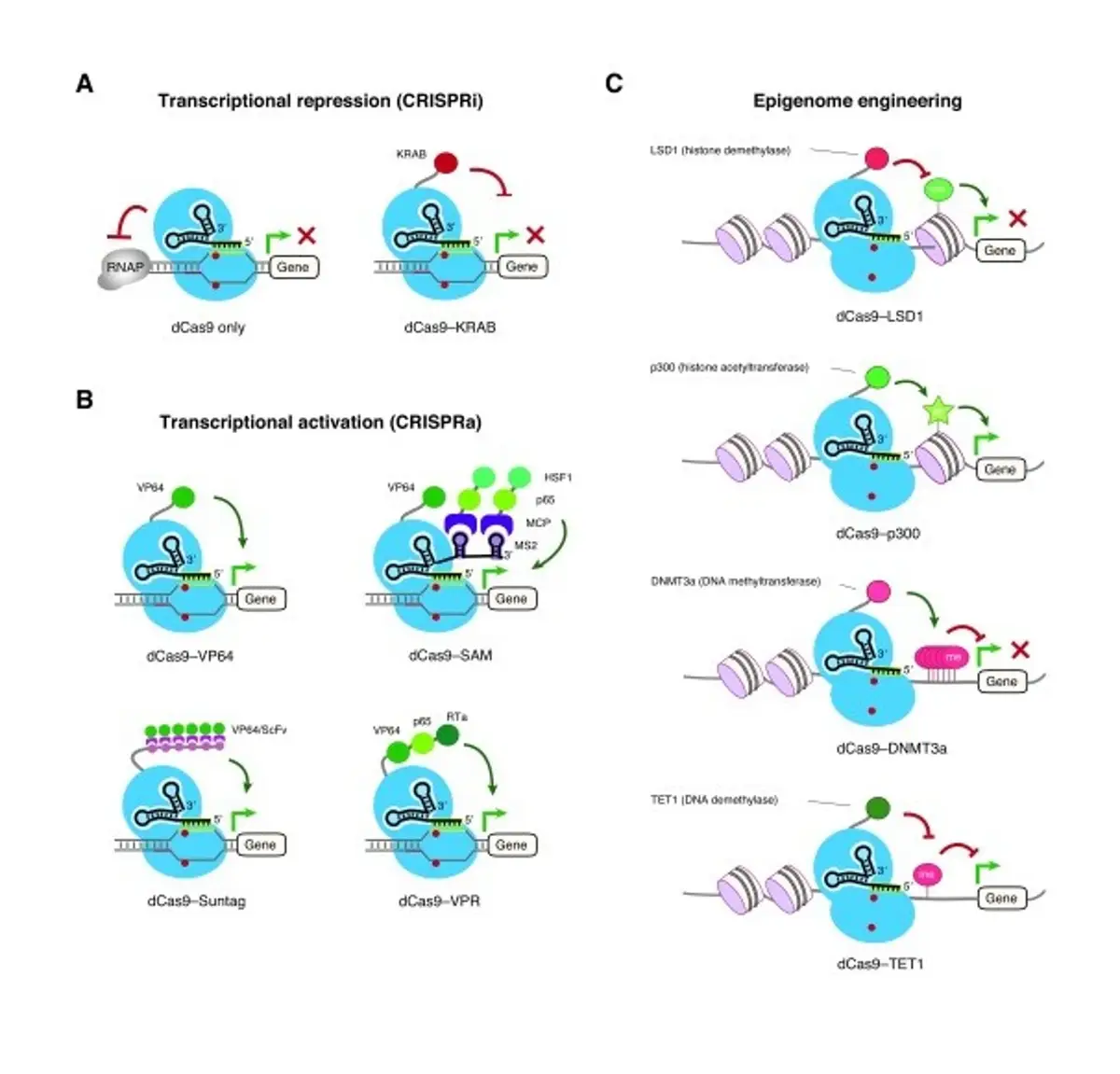
Image credit: Lo, A. & Qi, L. (2017). Genetic and epigenetic control of gene expression by CRISPR-Cas systems. F1000Res. 6.
A non-intuitive but useful application of CRISPR technology has been outside the realm of genome engineering. In 2013, Qi et al. created the dead Cas9 (dCas9) CRISPR variant by simply inactivating the catalytic nuclease domains while maintaining the recognition domains that allow guide RNA-mediated targeting to specific DNA sequences. The resulting nuclease-deactivated ‘dCas9’ was shown to silence gene expression by physically blocking the transcription.
When attached to effector domains like VP64, which is a transcriptional activation domain, these variants can be used to direct functional activity at any genomic region, either to activate or repress gene expression (CRISPRa and CRISPRi, respectively). Another development came when tagging a fluorescent dye to the dCas9 enabled the visualization of specific DNA fragments in the genome.
A second generation of dCas9 nucleases has since been developed, including dCas9-SAM, dCas9-SunTag, dCas9-VPR, and dCas9-KREB, each of which is fused to different effector domains. Of the CRISPRa nucleases, dCas9-SAM displayed the highest levels of gene activation. dCas9 nucleases have been used in many animal models and cell lines, including hPSCs, and can be used as epigenetic editors, with promise in cancer therapeutics. For a breakdown of how gene expression can be regulated with dCas9, you can watch this short video from ABM.
Experts have predicted that the future of genome engineering will involve gene writing, rather than gene editing. This hypothesis has already been proven correct with the establishment of base editing technology, which uses altered CRISPR systems to introduce single nucleotide variants (SNVs) to DNA or RNA. The first base editor enzymes consisted of a dead Cas9 domain fused with catalytic enzyme cytidine aminase that converted GC to AT. Researchers subsequently engineered a tRNA adenosine deaminase fused with Cas9 to convert AT to GC, thus allowing for a complete range of nucleotide exchanges in the genome. Due to the fact that roughly half of all known pathogenic genetic variants are caused by SNVs, base editing is an exciting avenue for precision medicine, with a recent explosion in base editing research by scientists who hope to use it to treat or cure genetic diseases. Since 99% of the SNVs in the human genome currently have unknown clinical implications, base editing technology will also assist in our understanding of how genetic variants affect human health. For more information, you can read this review on advances in base editing and potential applications of this technology in therapeutics.
In the years since CRISPR-Cas9 was discovered, we’ve found quite a few naturally occurring alternatives to Cas9 that have a wide variety of applications. In this section, we’ll take a look at the key Cas9 alternatives you should know about, and how they can be used. Around 50% of bacterial species and 90% of archaeal species are believed to possess CRISPR-Cas immune systems, so it’s likely that we are only seeing the tip of the iceberg when it comes to the different Cas nucleases that exist.
Although they might not be what you use for editing in your CRISPR experiments, we’d be remiss if we didn’t mention the Cas4-Cas1-Cas2 complex. It took researchers a while to figure out that this complex is actually a core component of CRISPR systems, with several studies attempting to elucidate the function of these nucleases.
Upon initial viral infection of a bacterial cell, the Cas1-Cas2 complex, which is highly conserved across all CRISPR-Cas systems, identifies and captures fragments of the foreign DNA (prespacers), and integrates them into the CRISPR array, providing immunity to future infection. The Cas4 has been found to be responsible for several key functions during this process in some (but not all) CRISPR systems. This includes trimming the fragments, ensuring they have the correct PAM sequence and cleaving the PAM, integrating the prespacers in the correct orientation, and preventing the premature integration of prespacers that have not yet been cleaved, which ensures the fidelity of spacers in the CRISPR array. These functions are crucial, allowing for successful CRISPR interference and survival of the host cell.
Cas3 differs significantly from other Cas nucleases. While Cas9 and its relatives will create clean, double-stranded breaks in DNA, Cas3 degrades DNA, moving along one strand and effectively ‘shredding’ it. It also lacks any PAM sequence requirements, being activated when the CRISPR-associated complex for antiviral defense (CASCADE) recognizes a target. Since it performs long-range, progressive, unidirectional DNA cleavage, Cas3 can effectively delete large regions (~10kb) of a target genome. This gives it the potential for many therapeutic targets, such as the removal of viruses that insert into host genomes and to elimination of antibiotic-resistant bacteria. The power of Cas3 is being utilized by biotechnology startup Locus Biosciences to engineer bacteriophage (known as crPhage), making them more efficient killers of bacteria for use in therapeutics. One such crPhage is currently being used in clinical trials to target Clostridioides difficile, a bacterial pathogen that causes urinary tract infections.
In 2016, Burstein et al. identified CasX and CasY from genome sequences of uncultivated bacteria from soil, water, biofilms, and other microbial communities. CasX and CasY are two of the most compact naturally occurring CRISPR variants and could have interesting applications as Cas9 substitutes in biotechnology. These nucleases also offer a glimpse into the potentially useful untapped nucleases from the vast microbial world.
Dr. Doudna and her colleagues published a study of the mechanism of action of CasX nucleases, concluding that they are a functionally distinct family. CasY is distinct because of its use of short-complementarity untranslated RNA (scoutRNA) together with crRNA, rather than tracrRNA like other CRISPR-Cas systems. CasX and CasY are now known by the names Cas12d and Cas12e, respectively.
Cas12a, first characterized by Zetsche et al., was originally called Cpf1, an abbreviation of ‘CRISPR from Prevotella and Francisella 1’. This nuclease has received considerable attention in the genome editing community in recent years because it has several interesting and useful features lacking in Cas9 variants. It has been used to edit a variety of targets in plant, bacterial, insect, and mammalian genomes.
As Cas9 requires the guanine-rich PAM sequence 5’ NGG 3’, it is not well suited for targeting AT-rich sequences. Cas12a, however, is well-suited for experiments targeting AT-rich DNA sequences. Cas12a nucleases from different bacterial sources recognize slightly different (but AT-rich) PAM sequences. While FnCas12a recognizes PAM sequence 5′‐TTN‐3′, AsCas12a and LbCas12a recognize 5′‐TTTV‐3′, where V is A, G, or C nucleotide.
One of the important features of Cas12a is that it creates staggered double-stranded breaks in the target DNA, rather than the blunt-end cuts generated by SpCas9. A key 2020 study by CRISPR co-inventor Dr. Jennifer Doudna and her colleagues at UC Berkeley elucidated the mechanisms by which Cas12a can generate ‘sticky end’ DSBs. It works by forming a 20 nucleotide, asymmetrical R loop, replacing one strand of the DNA double helix structure with the guide RNA, and placing the DNase active site for cleavage of the first strand. After the first cut, the DNA flanking the RNA-3’ side of the R loop becomes unstable, exposing the second strand of DNA for Cas12a to cut; the resulting overhang is five nucleotides in length.
Cas12a’s ability to create staggered-end DSBs makes it useful for experiments relying on the HDR repair outcome, like for CRISPR knock-ins to insert DNA into the genome. Also, Cas12a is smaller than SpCas9 and does not require a tracer RNA, only the crRNA. The gRNA required by Cas12a is, therefore, shorter in length, making it more economical to produce. This also has an advantage in the delivery of this CRISPR system, as it can be packaged into smaller delivery vehicles, like AAVs. It also has RNase activity, meaning that it is capable of processing its own guide RNA array and does not require tracrRNA. This makes it much more amenable to multiplexing experiments than Cas9.
Cas12a shows another interesting and useful trait; once it binds to and cleaves specific dsDNA, it is activated to indiscriminately and non-specifically cut any ssDNA. This ability of Cas12a can be exploited in diagnostics by using a reporter ssDNA, and it is being used for diagnostic applications in a number of studies Dr. Jennifer Doudna and her group have developed such as a method termed DNA Endonuclease Targeted CRISPR Trans Reporter (DETECTR). The team achieved high-sensitivity DNA detection of human papillomavirus in patient samples in proof of principle experiments. DETECTR has also been adapted for rapid Covid-19 diagnostics.
More recently, Cas12a was used by researchers at Harvard University’s Wyss Institute to create a wearable diagnostic device capable of detecting extremely low concentrations of the SARS-CoV-2 virus. The biosensor, which can be worn on clothing and face masks, is made up of lyophilized CRISPR enzymes loaded onto synthetic circuits and can be adapted to detect other pathogens by simply using different guide RNAs. For more information on the technology, you can read the full Nature Biotechnology paper on diagnostic face masks.
Importantly, Cas12a has been employed to edit the human genome for therapeutic purposes. A 2017 study, for example, used Cas12a to correct genetic mutations that cause Duchenne muscular dystrophy (DMD) in both mice and human cardiomyocytes generated from iPSCs. In 2021, Editas Medicine began the first clinical trial using Cas12a in human patients. The EDIT-301 treatment uses Cas12a to edit the beta-globin gene to successfully restore fetal hemoglobin in patient-derived hematopoietic stem cells (hPSCs) for the treatment of sickle cell disease.
This incredibly versatile Cas nuclease has been widely used to edit the genomes of various plants for agricultural purposes, including rice, soy, and tobacco. It has also been used to edit insect genomes, including the silk moth, Bombyx mori.
Similar to Cas9, Cas12a has also been engineered for a wider range of applications. A study published in Nature Biotechnology, described an enhanced variant from Acidaminococcus sp., enAsCas12a, which displayed an expanded target range of PAM sequences and significantly higher editing activity compared to wild type Cas12a. The researchers also engineered a high-fidelity variant, enAsCas12a-HF1 to reduce off-target editing. Delivered as ribonucleoprotein complexes, both new variants were tested for functionality in HEK293T and primary human T cells.
Various Cas12a nucleases are currently being used for diagnostic and gene therapy approaches by biotech startup Twelve Bio, which focuses specifically on Cas12a variants. This includes a diagnostic screening array for lung cancer, and a gene therapy tool repairing the mutations in the HBB gene that causes sickle cell anemia. For more details, you can read this interview with Stefano Stella, CEO of Twelve Bio, from CRISPR Medicine News.
Cas12b, formerly known as C2c1, is dual-RNA-guided, requiring both crRNA and tracrRNA to function. Characterized in Alicyclobacillus acidoterrestris (AacCas12b), this nuclease is generally smaller than its relatives Cas9 and Cas12a, which is an advantage. However, it functions at 48°C and therefore cannot be used in mammalian cells. To overcome this obstacle, Feng Zhang’s lab recently published a study of the Cas12b family and identified a variant that functions at 37°C, Bacillus hisashii (BhCas12b). Due to the lower efficiency of wild-type BhCas12b editing, they further created a mutant BhCas12b. The mutant demonstrated higher specificity than SpCas9, with promising results in editing human cell lines and primary T cells.
Cas12b variants that function at higher temperatures are still useful, however, with one becoming critical for detecting Covid-19 in a rapid diagnostic test. The adaptation of the SHERLOCK method, known as SHERLOCK testing, originally used Cas13a (described below in the Cas13a section). Feng Zhang’s lab wanted to create a more rapid test, with fewer fluid handling and incubation steps, but the sample needed to be incubated at 60°C. This temperature range is too hot for the Cas13a previously used, however after a screen of possible Cas nucleases, the team found Alicyclobacillus acidiphilus Cas12b (AapCas12b), which maintains activity at the right temperature for the test.
For more information on this technology, you can read our CRISPR Office Hours blog post on STOPCovid technology and listen to the CRISPR Cuts podcast episode, featuring Omar Abudayyeh and Jonathan Gootenberg.
Dr. Doudna reported compact Cas14 nuclease from archaea bacteria that demonstrated the following special features: it targets single-stranded (ss) DNA, does not require PAM sequence for activation, and cuts other ssDNA strands non-specifically upon binding the target sequence.
Although Cas14a is similar to the Cas12a nuclease in the aspect that it mediates cleavage of non-target ssDNA when activated by binding to a complementary DNA sequence, there is one main difference. Unlike Cas12a, Cas14a requires high-fidelity complementarity to the target ssDNA; it is particularly sensitive to mismatches in the internal seed region of the ssDNA target substrate. This quality makes it a prime candidate in diagnostics for recognizing single nucleotide polymorphisms in a genetic sequence.
All the nucleases described above target DNA, causing permanent changes in the genome. As RNA is transiently expressed, nucleases targeting RNA, rather than DNA, would be an excellent alternative to permanently modifying cells.
Nucleases targeting RNA were discovered in 2016 when Abudayyeh et al. characterized a Cas13a nuclease (known previously as C2c2) from the bacteria Leptotrichia shahii. The Cas13a nuclease was reported to contain two ribonuclease activities that separately catalyzed the maturation of CRISPR RNA (crRNA) and the degradation of single-stranded RNA in bacteria. The researchers, thereafter, also engineered Cas13a to target mRNA in mammalian cells.
Read our blog post on SHERLOCK CRISPR diagnostics.
Biochemical characterization of the enzyme revealed an interesting characteristic; the enzyme showed non-specific cleavage of any surrounding ssRNA after specific binding to the target site. Similar to Cas13a, Cas13b, which is also an RNA-guided RNAse, shows promiscuous ribonuclease activity on binding to its target.
Researchers realized the great potential of these Cas13 nucleases in diagnostics and used Cas13a in RNA detection in vitro. This method was further improved upon to achieve much higher sensitivity by researchers at the Broad Institute using their Specific High-sensitivity Enzymatic Reporter unLOCKing (SHERLOCK) tool.
In another application, researchers adapted dead Cas13 nucleases to edit RNA by directing an enzyme adenosine deaminase (which converts A to I) in mammalian cells. They called the system RNA Editing for Programmable A to I Replacement (REPAIR).
Two groups, led by Patrick Hsu from Salk Institute and David Scott from Arbor Biotechnologies, simultaneously discovered the Cas13d family of ribonucleases by scanning prokaryote sequences for nucleases resembling previously known Cas13 enzymes. These RNA-guided RNases are about 20% smaller than the Cas13a–Cas13c nucleases, but they show comparable targeting efficiency as the previously known variants. The smaller size of these enzymes gives them several advantages, such as being more convenient to package and deliver into cells.
Hsu and colleagues showed that CasRx, the best variant of their lot, had higher efficiency and specificity, compared to RNAi. They further targeted catalytically inactive CasRx not to cleave the RNA but to physically block access from the splicing factor, thus manipulating mRNA splicing in neuronal cells.
Cas13d has also been used to efficiently knock out genes involved in gene amplification, apoptosis, metabolism, and glycosylation in Chinese hamster ovary (CHO) cells, which are used for recombinant protein production in the biopharmaceutical industry. It has also been utilized in microbial RNA knockdown, gene silencing to study cancer progression, and knockdowns in several animal models.
CRISPR-Cas13d was also proposed as a treatment for Covid-19. The editorial, published in Cell Research, suggested that Cas13d could be used with gRNAs targeting the replicate-transcriptase (ORF1ab) and spike (S) regions of the virus, to cause functional disruption and limit its ability to replicate within human hosts.
Just like different tools are suitable for different functions, the choice of the nuclease to use in CRISPR experiments differs depending on the experimental goals. Altogether, at the current growth rate, we are progressing towards a future where a nuclease for every need will be available. Perfecting CRISPR technology and minimizing risks will open up the application of this method in humans.
Synthego's CRISPR nuclease portfolio includes the gold standard of gene editing, SpCas9, the novel high-fidelity Cas12Max, also referred to as hfCas12Max, and its newest nuclease engineered specifically for therapeutic applications, eSpOT-ON offered as recombinant protein and mRNA format. As your CRISPR Guide, we offer a full continuum of CRISPR solutions that can be used in various applications and enable the seamless development of your CRISPR-based therapeutics from the discovery to the clinic.
This protocol describes the use of limiting dilution to isolate single cells (clones) from a CRISPR edited population, the expansion of clonal populations, and screening to identify clones with the...
Thank you for choosing SpCas9 nuclease protein and gRNAs for your CRISPR experiment! SpCas9 has been used for diverse genome editing applications, making it the gold standard in CRISPR gene...
Learn more about clinical applications of CRISPR and how Synthego can support you from early-phase research, through process development, and into the clinic, highlighting our new sgRNA GMP manufacturing capabilities.